(перевод с английского)
Фотопродукты в ДНК, индуцированные солнечным светом
Спектры действия плоскоклеточной карциномы показывают, что ДНК является молекулой-мишенью; спектр поглощения ДНК хорошо коррелирует с летальностью, индукцией мутации и образованием фотопродуктов. 20- 24 Энергия , поглощенная ДНК производит молекулярные изменения, некоторые из которых включают одиночные базы, в результате чего другие взаимодействия между соседними и несмежных основаниями, а третьи между ДНК и белками. Относительные пропорции фотопродуктов ДНК будут варьироваться в УФ-спектре.
Димеризация между соседними пиримидинами - наиболее распространенные фотореакции, возникающие в результате прямого поглощения УФИ ДНК. Двумя основными фотопродуктами являются димер циклобутан-пиримидина (ЦПД) и, примерно на 25% частоты, фотопродукт [6-4] [(6-4) ПП] ( рис. 15.2 ). Распределение этих фотопродуктов в хроматине человека зависит от последовательности оснований, вторичной структуры ДНК и взаимодействий ДНК-белок. Поскольку цитозин более эффективно поглощает более длинные волны УФИ, чем тимин, CPD, содержащие это основание, легче образуются после облучения УФВ. 25 В сочетании с [6-4] PP, которые предпочтительно индуцируются тимин-цитозиновыми дипиримидинами, цитозиновые CPD могут играть главную роль в УФ-В (солнечном) мутагенезе. 26Последние данные показывают, что метилирование последовательностей PyrCG в гене p53 усиливает образование CPD в сайтах, которые являются горячими точками для мутаций. 27 PP может далее подвергаться UVB-зависимому превращению в его валентный фотоизомер, пиримидинон Дьюара 28 (см. Рис. 15.2 ). Помимо основных фотопродуктов, могут образовываться другие менее распространенные поражения, включая фотоаддукты пурин-пурин и пурин-пиримидин, фотогидратации и фотоокисления. 29 Поскольку общий выход этих фотопродуктов составляет всего 3-4% от выхода CPD, их биологическая роль считается минимальной; тем не менее, нельзя исключать их важность как премутагенные поражения на определенных участках.
![Рисунок 15.2. Фотохимические реакции в дипиримидиновой последовательности ДНК, приводящие к образованию CPD (TpT1, TpT2) или [6-4] PP (TpT4) и его фотолитического производного, пиримидона Дьюара (TpT3).](https://www.ncbi.nlm.nih.gov/books/NBK20854/bin/ch15f2.gif)
Рисунок 15.2
Фотохимические реакции в дипиримидиновой последовательности ДНК, приводящие к образованию CPD (TpT1, TpT2) или [6-4] PP (TpT4) и его фотолитического производного, пиримидона Дьюара (TpT3). (Перерисовано из Тейлора и Корса).
UVA в первую очередь вызывает повреждение косвенно через высокореактивные химические промежуточные соединения, кислородные и гидроксильные радикалы, которые, в свою очередь, реагируют с ДНК с образованием повреждений оснований, разрывов цепей и перекрестных связей ДНК-белок. Важность этих фотопродуктов неизвестна, но накапливаются доказательства того, что УФА может быть важным патогенным компонентом солнечного света. Значительные уровни гибели клеток и индукции мутаций наблюдались в эпидермальных клетках человека после облучения УФА-светом. 20, 22 Эти данные согласуются с более ранними исследованиями, которые предполагали, что летальные эффекты UVA не опосредованы повреждением CPD, 30, 31 и что поглотители свободных радикалов могут снизить цитотоксичность. 32Биологическое значение УФА света, возможно, лучше всего иллюстрируется недавней демонстрацией того, что УФА вызывает значительный уровень онкогенеза у лысых мышей. 33
Генетические факторы канцерогенеза кожи
Удаление УФ фотопродуктов
Идея о том, что УФ-повреждение ДНК является важным компонентом фотоканцерогенеза, возникла после открытия того, что клетки пациентов, страдающих наследственным заболеванием XP, испытывают дефицит в репарации ДНК. 34 Со времени этих первоначальных исследований молекулярная основа XP и родственных заболеваний в значительной степени решена, а основной механизм и соответствующие гены клонированы и охарактеризованы. 7, 35
Два основных пути эксцизионной репарации - эксцизионная репарация нуклеотидов (NER) и эксцизионная репарация оснований (BER) - воздействуют на различные виды повреждений. Путь NER, который связан с XP, удаляет пиримидиновые CPD и крупные химические аддукты в ДНК и заменяет поврежденный сайт вновь синтезированным полинуклеотидным участком длиной примерно 29 оснований ( рис. 15.3 ). 36, 37Путь BER удаляет основания ДНК, которые подверглись относительно небольшой степени модификации, такой как фотогидратация или фотоокисление. BER включает удаление поврежденного основания гликозилазами с последующим действием апуриновых / апиримидиновых эндонуклеаз плюс другие ферменты и кофакторы. Патч может быть меньше, чем результат NER; один путь вставляет только от одного до двух оснований, а второй второстепенный путь вставляет более крупный участок, близкий по размеру к участку NER. И NER, и BER представляют собой сложные процессы с участием нескольких генных продуктов, которые по-разному взаимодействуют с поврежденными участками в зависимости от точной химической формы и местоположения повреждения. Смежные основания, конформация ДНК, связанные белки,36
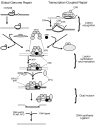
Рисунок 15.3
Биохимические этапы эксцизионной репарации нуклеотидов CPD в ДНК прокариот, показывающие биохимические детали событий, схематически представленных на рисунке 15.4. XPA связывается с фотопродуктами, и удаление происходит, когда УФ-специфические эндонуклеазы делают надрез
Эксцизионная репарация требует временной релаксации нуклеосомной структуры, так что поврежденные области становятся более доступными для экзогенных нуклеаз ( рис. 15.4 . Непрерывное удаление CPD и вставка оснований связано с очень низкой чистой частотой разрывов цепей ДНК, не более чем примерно 1 на 2 × 10 8 дальтон ДНК. Таким образом, только около 1% CPD, продуцируемых в ДНК дозой 10 Дж / м 2 , что представляет собой среднюю летальную дозу для клеток человека, подвергается иссечению в любой момент, но для восстановления одного поражения требуется всего около 4 минут. . Таким образом, эксцизия должна устанавливать динамический баланс между разрывом цепи и повторным соединением, и ее скорость ограничивается ферментами, участвующими в ранних стадиях репарации.

Рисунок 15.4.
Эвристическая схема эксцизионной репарации поврежденных участков ДНК в хроматине млекопитающих. Первый шаг включает механизмы, которые распознают повреждения и диссоциируют нуклеопротеины, чтобы сделать ДНК доступной для репарации ферментов. Далее следует последовательный разрез
Эксцизионная пластика - это неоднородный процесс. Между CPD и PP существует значительная разница в скорости их удаления из общего генома фибробластов и кожи грызунов и человека 38 даже несмотря на то, что основной механизм и размеры участков практически одинаковы. . 39 PP иссекаются быстрее, 50% удаляется из клеток человека и грызунов за 2-6 часов. CPD удаляются гораздо медленнее; половина удаляется из ДНК клетки человека в течение 12-24 часов, 40, 41но ничтожно малые количества удаляются из ДНК грызунов еще дольше. Частично удаление CPD может быть отложено из-за того, что сильное сродство системы вырезания к PP изначально блокирует доступные ферменты. Есть также большие различия в удалении CPD среди людей. Различная скорость иссечения может отражать тот факт, что PP значительно более искажают ДНК, и что они преимущественно расположены в межнуклеосомных областях ДНК, что может приводить к различиям в константе связывания между повреждениями - распознавание белков и ДНК. CPD распределены более хаотично, но с периодичностью 10Å в ДНК, обернутой вокруг нуклеосом, из-за предпочтения образования димеров на стороне ДНК, противоположной поверхности контакта ДНК-белок.
![Рисунок 15.5. Ремонт [6-4] PP и CPD в коже мышей и культивируемых клетках.](https://www.ncbi.nlm.nih.gov/books/NBK20854/bin/ch15f5.gif)
Рисунок 15.5.
Ремонт PP и CPD в коже мышей и культивируемых клетках. Радиоиммуноанализы, которые специфически обнаруживают PP ( A ) или CPD ( B ), использовались для мониторинга удаления этих повреждений из ДНК облученной кожи мыши (•) и мышиных клеток
![Рисунок 15.6. Ремонт [6-4] PP и CPD в человеческих фибробластах в культуре.](https://www.ncbi.nlm.nih.gov/books/NBK20854/bin/ch15f6.gif)
Рисунок 15.6.
Ремонт [6-4] PP и CPD в человеческих фибробластах в культуре. Радиоиммуноанализы, которые специфически обнаруживают PP или CPD, использовались для мониторинга удаления повреждений из ДНК нормального (GM637) или ревертанта XP
Когда удаление рассматривается на основе отдельного гена, существуют дополнительные вариации в соответствии с транскрипционной активностью. И NER, и BER происходят как глобальная репарация генома (GGR) и репарация, связанная с транскрипцией (TCR). CPD быстрее вырезаются из активно транскрибируемых генов, особенно из цепи ДНК, используемой в качестве матрицы для транскрипции. 44 Повышенная скорость удаления активных генов может также происходить для PP, но это нелегко разрешить на фоне большей общей скорости удаления этих фотопродуктов в геноме в целом. Различие в удалении активных и неактивных генов происходит потому, что основной фактор транскрипции, TFIIH, играет главную роль в репарации. 45Этот фактор регулирует базальную транскрипцию РНК-полимеразой II. Большинство генов, регулирующих TCR, связаны с заболеваниями человека XP, CS и TTD. Две из геликаз в TFIIH соответствуют генам XPB и XPD , а другие, как известно, играют роль своих аналогов в дрожжевом транскрипционном факторе b. 46 BER также может преимущественно протекать при транскрипции генов и, по-видимому, находится под влиянием таких разнообразных генных продуктов, как XPG , 47, ген репарации ошибочного спаривания MLH2 , 48 и ген предрасположенности к раку груди BRCA1 . 49 Детальное исследование промотора и первых экзонов PGKIген показал, что вырезание происходит медленно в областях связывания промотора, но увеличивается сразу после сайта начала транскрипции ATG. 50
Мутагенность УФ фотопродуктов
Два молекулярных механизма в настоящее время считаются важными в инициации канцерогенеза: активация протоонкогенов и инактивация генов-супрессоров опухолей. Оба участка действия уязвимы для летальных и мутагенных эффектов УФИ. Ген, такой как протоонкоген ras , может быть активирован точечной мутацией; p53, с другой стороны, представляет собой опухолевый супрессор, обычно инактивируемый точечными мутациями в опухолях человека. 51- 52 прогрессирования опухоли также зависят от УФРА. Смерть клеток из-за летального воздействия УФ-света может усилить клональную экспансию выживших клеток, которые могли быть мутированы или инициированы, увеличивая вероятность прогрессирования опухоли. 52Таким образом, взаимодействие УФ-летальности и мутагенеза в клетках кожи человека может определять начало и прогрессирование УФ-канцерогенеза, и супрессор опухолей p53 играет главную роль в этом определении. 52
Мутагенность и онкогенность конкретного фотопродукта может в конечном итоге зависеть от его летальности. Поражение, которое блокирует полимеризацию ДНК, может быть связано с прекращением синтеза ДНК. 51 Хотя большинство фотопродуктов действуют как блоки для репликативных полимераз, α и δ, их можно обойти во время синтеза ДНК в разной степени, в зависимости от их структуры, с помощью специфичных к повреждению полимераз η и ζ. 35 Этот обход позволяет полимеразам считывать неинформативную информацию о последовательности: полимераза η предпочтительно вставляет аденин в формирующуюся цепь напротив поражения (так называемое «правило А») и, следовательно, может точно воспроизводить тимин-содержащий CPD, 53, 54, тогда как полимераза ζ мутагенный. 55Этот механизм имеет два важных значения, касающихся мутагенности различных фотопродуктов. Во-первых, мутации чаще всего возникают там, где цитозин является компонентом фотопродукта, поскольку вставка аденина напротив тимина является правильным и немутагенным событием. Следовательно, большинство CPD, поскольку они образуются между двумя основаниями тимина, не являются мутагенными. Во-вторых, чем более искажено поражение, тем больше вероятность, что оно заблокирует синтез ДНК и приведет к летальному исходу, а не к мутагенному событию. Поскольку ] PP значительно более деформирует, чем CPD (т. Е. Вызывает изгиб спирали на 47 °, а не на 7 °), он с большей вероятностью будет летальным, чем мутагенным. Поскольку обходное повреждение и введение аденина зависят от множества условий, как CPD, так и PP вносят сложный вклад в онкогенез.
Сайт-специфическое определение индукции фотопродуктов в гене lacI Escherichia coli предполагает корреляцию между горячими точками индукции PP и УФ-индуцированных мутаций. 56 Анализ сайтов индукции PP показал, что это поражение является ответственным за большую часть мутаций перехода цитозина в тимин в E. coli . В соответствии с этим наблюдением было показано, что исключительная индукция CPD ацетофеноном и УФ-светом не увеличивает индукцию переходных мутаций в фаге лямбда. 57 Подобная взаимосвязь наблюдалась при исследовании фотореактивации в E. coli; в то время как CPD и PP были одинаково цитотоксичными, последние были гораздо более мутагенными. 58
Роль определенных фотопродуктов в УФ-мутагенезе в клетках человека была исследована с использованием челночных векторов. В этих системах плазмиды на основе обезьяньего вируса, облученного ультрафиолетом (SV40), трансфицируются в клетки человека, где они реплицируются хозяином. Затем плазмиды выделяют, амплифицируют в бактериях и анализируют на предмет индукции мутации путем секвенирования ДНК. Затем сайты мутаций можно сравнить с сайтами индукции фотопродукта в целевой последовательности. Результаты этих исследований аналогичны результатам, полученным на E. coli : сайты переходных мутаций коррелируют с сайтами повышенной индукции PP ( таблица 15.1 ). В частности, сайты и частоты горячих точек мутаций в lacIгены, трансфицированные в клетки человека, были идентичны генам, определенным в E. coli . 59 В системе челночных векторов, в которой индукция фотопродукта и сайты мутации были исследованы в гене supF , трансфекция в SV40-трансформированные человеческие фибробласты и клетки почек обезьяны показала аналогичную корреляцию. 60 В гене supF , встроенном в хромосому L-клеток мыши 61 и в эндогенном гене APRT клеток CHO, 62 большинство мутаций состояло из переходов цитозин-тимин, происходящих в последовательностях тимин-цитозин и цитозин-цитозин. Из-за специфичности репарации цепи существует смещение между мутациями в кодирующей и некодирующей цепях экспрессируемых генов, которое различается в зависимости от способности клеток к NER. 63, 64
Таблица 15.1
UVC-индуцированные мутации, наблюдаемые в челночном векторе pZ189, реплицированном в XP или нормальных клетках человека а .
И CPD, и PP могут образовываться в последовательностях, которые, как было показано, являются горячими точками мутаций в челночных векторах, и идентичность мутагенного поражения была проверена фотореактивацией последовательности supF в плазмидах перед трансфекцией. 65, 66 Ферментативное фотореверсирование CPD снижает частоту мутаций в нормальных клетках на 75% и в клетках XP группы A на 90%. Поскольку котрансфекция обезьяньих клеток смесью необлученной плазмиды supF и облученной плазмиды без гена supF не вызывала мутаций, роль SOS-подобной системы, наблюдаемая в E. coli , по-видимому, не является ответственной за полученные результаты. 66 Эти результаты не согласуются с моделью, разработанной вE. coli и предполагают, что PP могут быть менее мутагенными в человеческих клетках. Аналогичный анализ с фотореактивацией показал, что CPD, встречающиеся в дипиримидиновых сайтах, содержащих по крайней мере одно цитозиновое основание, были преобладающими мутагенными повреждениями, индуцированными в клетках человека, и что [6-4] PP на этих сайтах составляли только около 10% мутаций. 65 Однако это же исследование показало, что частоты как CPD, так и [6-4] PP в отдельных сайтах дипиримидина не коррелировали с частотой мутаций, предполагая, что, хотя для мутагенеза необходимы УФ-индуцированные поражения, горячие точки мутаций определяются другими факторами. .
Однако сравнение выходов фотопродуктов, скорости репарации и мутаций в генах PGKI и p53 показало, что области с высоким уровнем УФ-индуцированных мутаций могут быть вызваны высоким выходом фотопродуктов и / или низким уровнем репарации. 50, 67- 69 Комбинации исходных урожайности и темпов ремонта , которые оставляют высокую чистую постоянную нагрузку фотопродуктов в конкретном месте , как представляется, непосредственно связана с мутационным выходом. Используя опосредованную лигированием полимеразную цепную реакцию (LMPCR), которая позволяет точно локализовать поврежденные основания, было картировано распределение фотопродуктов в экзонах 1 и 2 трех протоонкогенов ras , и не было корреляции между частотой фотопродуктов и индукцией мутации в кодоне 12 H - рас и К-ras был найден. 70 Дальнейшие исследования с LMPCR показали, что скорость эксцизионной репарации CPD на специфических нуклеотидах в промоторе и экзоне 1 гена PGK1 варьировала в 15 раз при значительном снижении репарации в сайтах связывания факторов транскрипции. 50 Восстановление ДНК отдельных нуклеотидов в гене-супрессоре опухоли p53 было сильно вариабельным и зависимым от последовательности, с медленным восстановлением, наблюдаемым в семи из восьми положений, связанных с мутациями. 67 УФ-индуцированные мутации в гене p53 являются вероятной ступенью в формировании плоскоклеточной карциномы 51, 52 и могут возникать в «холодных точках» репарации ДНК, а не в «горячих точках» фотопродуктов.
Генетические нарушения восстановления ДНК
Изучение заболеваний, чувствительных к солнечному свету, и отбор клеток хомяков и мышей, чувствительных к ультрафиолету, в культуре выявили большой ряд генетических локусов, которые контролируют реакцию кожи млекопитающих на повреждение ( таблица 15.2 ). Все эти локусы характеризуются значительным повышением чувствительности к УФ-излучению или УФ-В излучению и включают нарушения XP (8 групп комплементации), 7 CS (2 группы комплементации), 71, 72 TTD (3 различных фенотипических типа), 73, 74 и BCNS. . 75 Многие из этих генов также были определены в культурах клеток как ERCC1-12 (эксцизионная репарация перекрестного комплемента). 76
Таблица 15.2
Группы комплементации в XP и УФ-чувствительных клетках яичника китайского хомячка (CHO).
За исключением BCNS, все эти нарушения представляют собой повышенную чувствительность к длинам волн UVB и UVC из-за рецессивных мутаций, связанных с большим семейством генов, которые регулируют репарацию ДНК клеток человека. Эти нарушения не исключают друг друга, поскольку CS перекрывается с группами XP B, D и G, а TTD перекрывается с группами B и D. Расположение хромосом известно, и гены были клонированы для большинства этих локусов (см. Таблицу 15.2 ). Мутации в отдельных генах оказывают плейотропное действие на чувствительность клеток к ультрафиолетовому излучению и восстановление ДНК и связаны с рядом клинических синдромов, затрагивающих кожу, нервную систему и иммунологические изменения. BCNS, напротив, является доминантным заболеванием, включающим мутацию в супрессоре опухоли, который регулирует сигнальный каскад, включающий исправленный (PTC ) ген. 77
Механизм эксцизионного восстановления нуклеотидов
Пиримидиновые CPD и PP, продуцируемые в ДНК ультрафиолетовым или ультрафиолетовым излучением B, восстанавливаются с помощью сложного многоступенчатого процесса, включающего множество взаимодействующих генных продуктов. Отчасти именно потребность во взаимодействующих белках в процессе репарации приводит к возникновению сложных перекрывающихся симптомов у некоторых пациентов с мутациями в этих генах. Процесс репарации, в принципе, включает удаление олигонуклеотида из 27–29 нуклеотидов, содержащего фотопродукт, путем точно расположенных расщеплений 5 нуклеотидов на 3'-стороне фотопродукта и 24 нуклеотида на 5'-стороне. 78 После удаления этого олигонуклеотида образовавшаяся брешь заполняется ДНК-полимеразой δ, ядерным антигеном пролиферирующих клеток (PCNA) и одноцепочечным связывающим белком и лигазой. 36Эти процессы можно рассматривать как включающие последовательные этапы распознавания фотопродукта, сборки эксцизионного комплекса, смещения вырезанного фрагмента и полимеризации замещающего пластыря.
Распознавание фотопродуктов достигается за счет специфического связывания и ассоциации нескольких белков. Продукт гена XPA был первым идентифицированным белком распознавания повреждений и, по-видимому, ограничивает скорость восстановления в клетках человека. 79 Распознавание может происходить из-за искажений и одноцепочечной ДНК в фотопродуктах, или фотопродукты могут вылетать из спирали ДНК в карман в белке, как это происходит в некоторых других ферментах репарации ДНК. 80 XPC- hHR23B комплекс 81, 82 является самым ранним детектором повреждений , чтобы инициировать NER в нетранскрибируемой ДНК, действуя до белка РФА, и служит для стабилизации РФА связывания с поврежденным сайтом с высоким сродством к] PP. 83, 84 Белок XPC может потребоваться для временного разворачивания нуклеосом во время NER. 85 Этот комплекс специфически участвует в GGR, но не в TCR, где блокировка РНК-полимеразы II на поврежденном основании может функционировать на его месте. Стабильная ассоциация TFIIH с повреждениями ДНК зависит от целостности белков XPA и XPC. Помимо белков XPA и XPC, белок XPE имеет сходные характеристики связывания, но играет гораздо менее заметную роль. Две субъединицы, которые совместно очищаются, связаны с XPE: p48, который, как обнаружено, несет мутации от нескольких пациентов с XPE, и белок p125, и они могут участвовать в репарации менее доступных повреждений в нетранскрибируемой ДНК. 86 Субъединица p48 индуцируется в клетках человека и не экспрессируется в клетках хомяка, которые не могут восстанавливать CPD в нетранскрибируемой ДНК. Уровень мРНК p48 сильно зависит от базовой экспрессии p53 и может обеспечивать связь между p53 и NER. 87
NER действует скорее за счет сборки отдельных факторов в местах повреждения ДНК, чем за счет предварительной сборки голокомплексов. 88 Основные белковые факторы включают белок XPA, гетеротримерный белок репликации (RPA), субъединицу TFIIH с 6 по 9, комплекс XPC- hHR23B , нуклеазу XPG и нуклеазу ERCC1 -XPF. 36 После сборки комплекс XPC- hHR23B диссоциирует, и белок XPG отрезает 3 'от поражения и ERCC1-Гетеродимер -XPF отрезает 5 'от CPD. Комплекс нуклеазы плюс однонитевой фрагмент длиной 29–30 нуклеотидов высвобождается под действием фактора транскрипции TFIIH, который содержит как 3'-5 '(XPB), так и 5'-3' (XPD) геликазы. Белок XPG также необходим для TCR окислительного повреждения. 47 По крайней мере, один компонент TFIIH, XPB, взаимодействует с p53 и инициирует сигнальный каскад, ведущий к апоптозу в поврежденных клетках. 89 Для работы всего процесса NER требуется около 100 нуклеотидов ДНК. 78 PCNA, которая необходима для репарационного синтеза, также взаимодействует с GADD45, белком, индуцируемым повреждением, который стимулирует эксцизионную репарацию in vitro, хотя его функция in vivo неизвестна. 90 Многие компоненты всего механизма эксцизионной репарации являются продуктами генов, которые вызывают различные чувствительные к солнцу расстройства и нарушения развития. Некоторые компоненты эксцизионной репарации, особенно гены ERCC1 91, 92 и hHR23B 82 , не были обнаружены в группах дефектной эксцизионной репарации комплементации. Инактивация гена ERCC1 приводит к образованию чувствительных к УФ-излучению клеток и вызывает летальную печеночную недостаточность у мышей. 91 Сложность этих заболеваний проистекает не только из особого порядка, в котором генные продукты играют в репарации, но также из-за вторых ролей в факторах транскрипции и сигнальных каскадах.
Пигментная ксеродермия
XP - это редкое аутосомно-рецессивное заболевание, которое встречается в США с частотой 1: 250 000. 7Пораженные пациенты (гомозиготы) имеют чувствительность к солнцу, что приводит к прогрессирующим дегенеративным изменениям подверженных воздействию солнца участков кожи и глаз, часто приводящим к неоплазии. У некоторых пациентов XP, кроме того, прогрессирующая неврологическая дегенерация. Облигатные гетерозиготы (родители) обычно протекают бессимптомно. Средний возраст начала заболевания составляет от 1 до 2 лет, при этом кожа быстро приобретает вид, который наблюдается у людей, много лет находившихся на солнце. Пигментация неоднородна, на коже наблюдаются атрофия и телеангиэктазия с развитием базальноклеточного и плоскоклеточного рака. Частота онкологических заболеваний примерно в 2000 раз выше, чем у населения в возрасте до 20 лет, при этом продолжительность жизни сокращается примерно на 30 лет.
Клетки от пациентов с XP удаляют CPD пиримидина и [6-4] PP со сниженной скоростью от 0 до 90% от нормы, за исключением группы вариантов, которая имеет показатели, близкие к нормальным ( Таблица 15.3 ). Уменьшение иссечения коррелирует с низким уровнем репликации репарации. Снижение сходно во всех исследованных тканях, включая кожу in vivo, периферические лимфоциты, фибробласты, культуры клеток печени и опухолевые клетки. Среди пациентов с дефицитом эксцизионной репарации существует семь групп комплементации, а восьмая, вариант XP, имеет дефект репликации поврежденной ДНК (см. Таблицу 15.2 ).
Таблица 15.3
Характеристики репарации ДНК человеческих клеток.
Внутри этих заболеваний существует значительное генетическое разнообразие, и способность к эксцизионной репарации во многих случаях коррелирует со способностью выживать при УФ-облучении. По сравнению с нормальными клетками, клетки из XP групп A и D очень чувствительны к летальному воздействию ультрафиолетового света и неспособны исключить два основных типа ультрафиолетовых повреждений, CPD и [6-4] PP (см. Таблицу 15.3 ). . Клетки группы А XP также обладают пониженной способностью восстанавливать пиримидинон Дьюара, важное повреждение, индуцированное с повышенной эффективностью под действием УФ-В-света. 26, 38, 93 Клетки группы Е XP демонстрируют промежуточный фенотип как по своей чувствительности к УФ-излучению, так и по способности устранять УФ-повреждения, а ряд XPE-клеток лишен связывающего белка, специфичного к повреждению p48 / p125. 94
Группа C является одной из самых больших групп и часто упоминается как обычная или классическая форма XP. У пациентов наблюдаются только кожные заболевания, которые значительно различаются по степени тяжести в зависимости от климата. Опухоли языка наблюдались у нескольких пациентов. Клетки имеют низкие, но неоднородные уровни эксцизионной репарации (10–20% от нормы) и менее чувствительны к уничтожению УФ-светом и химическими канцерогенами, чем клетки в группах A и D. Одной из характеристик репарации, уникальной для этой группы, является то, что сниженная репарация не широко распространен в их геноме, например, в группах A и D, но ограничен определенными областями генома. 95 Эти клетки вставляют участки репарации в небольшие участки своего генома с нормальной скоростью, что, вероятно, соответствует репарации [6-4] PP и CPD 95и вырезать CPD тимина предпочтительно из транскрипционно активных областей. 95 Это ставит дилемму о том, что высокие уровни гибели клеток, соматических мутаций и рака из-за УФ-излучения в XP-группе C связаны с дефицитом репарации в нетранскрибируемых областях генома. Это, в свою очередь, предполагает, что активирующие, а не заглушающие мутации могут быть важны, или что мутации возникают из-за нерепарированных повреждений в нетранскрибируемой цепи активных генов.
Группа E - это редкая группа, которая проявляет легкие симптомы и остаточные уровни восстановления, которые составляют от 50 до 100% от нормы. Некоторые клетки XPE не имеют ДНК-связывающего белка 94 и, соответственно, несут мутации в гене p48, но это наблюдается не во всех случаях, что повышает вероятность того, что не все клетки, классифицированные в группе E, правильно отнесены. Роль этого белка все еще неясна, но он зависит от р53 и участвует в репарации нетранскрибируемых участков ДНК. 86, 87
Клетки варианта XP не имеют дефекта эксцизионной репарации, но кажутся близкими к нормальным по своей способности вырезать повреждения ДНК (см. Таблицу 15.3 ), но демонстрируют небольшую, но значительную чувствительность к ультрафиолетовому свету. Теперь известно, что в этой группе комплементации отсутствует ДНК-полимераза, HRAD30 (полимераза η), которая необходима для точной репликации димеров пиримидина. 96, 97В его отсутствие клетки варианта XP очень восприимчивы к УФ-индуцированному мутагенезу и связаны по существу с теми же симптомами, что и другие пациенты XP. Канцерогенез в результате УФ-повреждения у пациентов с XP возникает, следовательно, из-за потери либо способности NER, либо полимеразы η; оба приводят к увеличению количества повреждений ДНК, которые становятся субстратом для генерации ошибок (таких как мутации, перестройки генов и делеции) полимеразой ζ.
Синдром Кокейна
CS - аутосомно-рецессивное заболевание, характеризующееся кахектической карликовостью, аномалиями сетчатки, микроцефалией, глухотой, нервными дефектами и задержкой роста и развития после рождения. Карциномы кожи в результате гиперфоточувствительности не наблюдаются у пациентов с CS, что отличает это заболевание от XP.
Пациенты с CS распределены неравномерно в группах комплементации: в группе A значительно больше пациентов, чем в группе B. 71 Из группы B дополнения XP известны три пациента из двух семей, у которых также проявляются симптомы CS. 98 симптомов CS также были зарегистрированы у нескольких пациентов с XPD и XPG. Чувствительность к УФ-излучению большинства клеток CS находится в узком диапазоне, с D 37 примерно наполовину от нормы, в отличие от клеток XP, которые демонстрируют широкий диапазон чувствительности. Характерные клеточные изменения CS включают неспособность синтеза ДНК и РНК вернуться к нормальным уровням после УФ-облучения. 72 Удаление фотопродуктов ДНК из тотальной геномной ДНК клеток CS является нормальным явлением, но восстановление транскрипционно активных генов снижено. 99 Продукты гена CS участвуют в сопряжении эксцизионной репарации с транскрипцией, но их точная функция еще не ясна. Они могут быть вовлечены в убиквитинирование и деградацию застопорившейся РНК polII на поврежденных участках.
Таким образом, синдром Кокейна и группа C XP составляют интересный контраст. Клетки CS восстанавливают только транскрипционно неактивные гены, тогда как клетки XP группы C восстанавливают только транскрипционно активные гены. Они демонстрируют аналогичную повышенную чувствительность к уничтожению клеток, указывая на то, что все области генома должны быть восстановлены для нормального выживания. Но только XP группа C демонстрирует повышенный мутагенез и канцерогенез, что указывает на то, что дефектная репарация транскрипционно неактивных генов более важна для этих конечных точек.
Трихотиодистрофия
TTD - редкое аутосомно-рецессивное заболевание, характеризующееся хрупкостью волос с дефицитом серы и ихтиозом. Стержни волос продольно расщепляются на мелкие волокна, и эта ломкость связана с уровнями цистеина / цистина в белках волос, которые составляют от 15 до 50% от таковых у нормальных людей. Состояние также сопровождается задержкой физического и умственного развития различной степени тяжести. Пациенты часто имеют необычный внешний вид лица с выступающими ушами и опущенным подбородком. Умственные способности варьируются от низко-нормальных до тяжелой отсталости. 73 На основе клеточных реакций на УФ-повреждение можно выделить три категории заболевания. Наиболее серьезные из них имеют дефекты ремонта и дополняющие свойства, которые помещают их в группу D по опыту; в промежуточных случаях наблюдается пониженная репарация ДНК, но нормальная чувствительность к ультрафиолету; а третий по УФ-ответу неотличим от нормальных клеток. 74
Были охарактеризованы профили восстановления трех линий фибробластов, полученных от пациентов с TTD; каждый демонстрирует уникальный фенотип. 74 Одна линия клеток TTD показывает нормальную устойчивость к ультрафиолету и способность к репарации ДНК; другой показывает ответ XP группы D на УФ-облучение со значительным снижением выживаемости и репарации и связан с мутациями в гене XPD; 100 клетки, полученные от третьего пациента, показывают нормальную выживаемость после УФ-облучения, но способность к восстановлению, о чем свидетельствует синтез восстановления и ремонтный разрез, значительно снижается. Хотя скорость иссечения [6-4] PP в этом третьем классе TTD немного снижена, восстановление CPD кажется нормальным, что позволяет предположить, что либо PP не является летальным для этих клеток, либо наблюдаемый дефект в его репарации проявляется. недостаточно, чтобы повлиять на выживание. Конкретные гены TTD (A, B) могут быть компонентами фактора транскрипции TFIIH, 46 и симптомы этого заболевания указывают на роль этого фактора в развитии и росте волос.
Синдром базально-клеточного невуса и базально-клеточный рак
BCNS - аутосомно-доминантное заболевание с высокой пенетрантностью (> 97%). Основными проявлениями этого синдрома являются множественные опухоли (в среднем от 50 до 100), в основном на коже, подвергающейся воздействию солнечных лучей, которые обычно появляются в период полового созревания, а также во втором и третьем десятилетии жизни. 75 Другие симптомы включают ямки на ладонях и подошвах, а также аномалии опорно-двигательного аппарата (сколиоз, раздвоение ребер, расщепление позвоночника). Высокая частота аномалий развития предполагает, что нормальный аллель гена BCNS может играть роль в росте и развитии, помимо ускорения канцерогенеза, вызванного солнечным светом. Ген, связанный с BCNS, был идентифицирован как ген, ранее идентифицированный у Drosophila как патчированный ген ( PTC ). 77 Мутации вPTC были идентифицированы при спорадических базальных клетках рака (BCC) и у пациентов с BCNS, а также с повышенной частотой в BCC у пациентов XP. 101
Наиболее частым грубым генетическим изменением при спорадических BCC (68%) является потеря гетерозиготности (LOH) в локусе PTC на хромосоме 9q22. 102 Это происходит даже в небольших BCC (<1 см в диаметре), во время их начального развития из стволовых клеток волосяных фолликулов, предполагая, что LOH в локусе PTC может быть ранним событием в онкогенезе BCC. 102 BCNS-ассоциированные BCCs сохраняют мутантный аллель PTC зародышевой линии, но теряют аллель дикого типа (WT) из-за потери больших хромосомных фрагментов на 9q (Bonifas et al, 1994) или из-за точечной мутации. В соответствии с гипотезой о двух ударах Кнудсона, у пациентов с BCNS развивается больше BCC, от десятков до сотен и в более молодом среднем возрасте, чем в большинстве спорадических случаев.
Ген PTC человека кодирует мембранный белок из 1296 аминокислот, который, как предполагается, имеет 12 трансмембранных доменов, которые регулируют клеточную пролиферацию и путь передачи сигнала. Белок PTC подавляет экспрессию гена hedgehog (HH), взаимодействуя с семи трансмембранным белком, напоминающим рецепторный белок, связанный с G-белком (Smoothened, SMO). PTC- ингибирование SMO снимается связыванием PTC с SHH или последующей мутационной инактивацией PTC . 77 Передача сигналов SMO может активировать транскрипцию мишеней HH, включая PTC, через сигнальный путь, ведущий к активации фактора транскрипции, Gli. Таким образом, как это ни парадоксально, мутационная инактивация PTC и, как следствие, потеря активности белка PTC приводит к увеличениюЭкспрессия PTC и накопление высоких уровней транскрипта PTC .
Развитие других опухолей в BCNS, включая медуллобластому и фибромы яичников и матки, предполагает, что BCNS соответствует модели двух мутаций Кнудсона для канцерогенеза. 75 Однако количество таких мутаций, необходимых для индукции рака у людей с BCNS, неизвестно и может быть больше двух. Тем не менее, в BCNS одна из мутаций наследуется как аутосомно-доминантный ген во всех соматических клетках, тогда как факторы окружающей среды, такие как УФ или ионизирующее излучение, могут вызывать оставшуюся мутацию (и). Эта гипотеза подтверждается наблюдением, что у несимптомных детей с BCNS, получавших лучевую терапию по поводу медуллобластомы, развились множественные базально-клеточные карциномы в области, подвергшейся облучению от 6 месяцев до 3 лет спустя.
Фибробласты от людей с BCNS не показали последовательного повышения чувствительности к рентгеновским лучам или УФ-излучению. Хотя восстановление CPD в этих клетках является нормальным, удаление [6-4] PPs может быть уменьшено, напоминая то, что показано для TTD группы 3 пациентов 103 (Rosenstein B, Mitchell DL неопубликованные наблюдения). Сниженная репарация [6-4] PP может быть недостаточной, чтобы повлиять на выживаемость после УФ-облучения, и в целом BCNS, по-видимому, не имеет серьезных отклонений в УФ-репарации.
Семейная меланома
Меланома вызывается воздействием солнца, но точный механизм гораздо менее ясен, чем при немеланомном раке кожи. 4, 104 Заболеваемость меланомой увеличивается у пациентов с XP, что указывает на то, что УФ-облучение и повреждение ДНК могут быть вовлечены в индукцию меланомы. 9 Но индукция кажется более тесно связанной с острыми ожогами, чем с хроническим накопленным воздействием. Семейные случаи указывают на то, что на хромосоме 9q21 находится главный супрессор опухоли меланомы, который кодирует регулятор клеточного цикла, p16, ингибитор семейства генов циклинкиназы 4. 104 Потери генов и гетерозиготность в этой хромосомной области наблюдаются как в семейных, так и в несемейных случаях, что демонстрирует их общую важность. 105Также задействованы дополнительные гены на хромосомах 6, 8 и 10. 105, 106 Делеция хромосомы 9p21 в значительном количестве случаев может распространяться на дополнительную потерю на больших участках 9p и 9q. 105 Так как плечо 9q также содержит гены BCNS и XPA, эти дополнительные потери, вероятно, могут вносить вклад в изменения светочувствительности и репарации, связанные с прогрессированием меланомы.
Канцерогенез
Канцерогенез часто, кажется, протекает в многоэтапном процессе, первый из которых является инициирующим событием с последующими рекламными мероприятиями, которые часто могут происходить намного позже. Согласно одной из точек зрения на канцерогенез, инициация коррелирует с индукцией соматических мутаций, а продвижение - с дальнейшими изменениями экспрессии генов и количества копий. Канцерогенез, по-видимому, связан с активностью большого количества генов. К ним относятся гены детоксикации канцерогенных химических веществ, семейство генов репарации ДНК, около 50 или более доминантно действующих протоонкогенов, активируемых мутацией, делецией, транслокацией или амплификацией, а также гены-супрессоры опухолей, потеря которых может способствовать развитию рака. 107, 108
Последовательность событий, наблюдаемых при колоректальном раке и ретинобластоме, может служить полезной моделью канцерогенеза кожи. 107, 108 Ранние события могут соответствовать активирующим мутациям, и различные стадии развития опухоли возникают в результате прогрессирующей потери хромосом или преобразования гетерозиготности в гомозиготность. На основании исследований с XP, ранние события на коже могут соответствовать УФ-индуцированным мутациям. Мало того, что основные генетические дефекты репарации связаны с раком при XP, но и вариации репарации у разных людей также показывают корреляцию с базальноклеточной карциномой. Однако здесь недавние исследования активации ras привели к дилемме. Несколько исследований привели к идентификации активирующих мутаций в Ha- ras и N-ras онкогены в кодоне 61, от солнечного УФ-облучения. 109 ~ 111 Однако, несмотря на более чем 75% от УФ-индуцированных мутаций С до Т переходов на ТС или сайты фотопродукта CC НПР, 112 HA- РАН и N- Ras активации произошло в опухолях на ТТ - сайте и трансверсии ранее не идентифицированы в модельные культурные системы. 112 Очевидно, что необходимо подробное исследование активации онкогенов в ряде систем мыши и человека, чтобы прояснить взаимосвязь между УФ-индуцированными мутациями и активацией ras . Большая часть опухолей кожи человека содержит мутации в гене-супрессоре опухолей p53, которые вызваны УФ-фотопродуктами. 51- 52Это демонстрирует прямую причинную роль УФВ от солнечного света в возникновении одного из мутагенных явлений при канцерогенезе кожи и демонстрирует, что мутации р53 являются ранними событиями, которые влияют на баланс путей гибели клеток по сравнению с мутациями и пролиферацией.
Инактивация генов-супрессоров опухолей была продемонстрирована в ретинобластоме 113, опухоли Вильмса 114 и акустических невриномах, и потеря аллелей, приводящая к преобразованию из гетерозиготности в гомозиготность, по-видимому, является частым следствием прогрессирования опухоли. 108 Высокие уровни рака кожи у пациентов с XP могут быть результатом повышенного уровня УФ-повреждения, вызванного дефектной репарацией, что приводит к активирующим мутациям, инактивации мутаций p53 и нестабильности хромосом. Гены-супрессоры опухоли могут способствовать развитию опухоли за счет дополнительных эффектов на рост клеток и межклеточную регуляцию, а также потерю пути апоптоза из-за гетерозиготности p53. 51, 52Наблюдение, что продвижение может включать изменения в коммуникации между ячейками, согласуется с этой интерпретацией. Промоторы опухоли могут быть факторами окружающей среды, имитирующими действие регуляторных генов. Анализ различных стадий развития кожной опухоли может показаться особенно многообещающим в настоящее время, поскольку доступно так много стадий и так хорошо известны причинные факторы окружающей среды. Значительное сокращение времени индукции рака у пациентов с XP (см. Рис. 15.1 ) указывает на то, что УФ-повреждение и восстановление участвуют как в инициировании, так и в прогрессировании рака кожи и, следовательно, являются критическими факторами на протяжении всего канцерогенного процесса.
